
光受容タンパク質とオプトジェネティクス

この記事では、眼の網膜に存在する光受容タンパク質と、オプトジェネティクス(光遺伝学)についてまとめます。
ものが見えるしくみ

ものがみえるしくみは、カメラの構造に例えられる。
眼球の前方にある角膜(レンズ)で取り入れた光は、その内側の水晶体(遠近調節レンズ)を通過して、奥のフィルム(網膜)に投射する。
網膜には、1億個以上の視細胞が集まっている。
視細胞で受容した光は、電気シグナルに変換される。そして電気シグナルは視神経を通して脳へと伝えられ、脳で処理された情報をはじめて「見る」ことができる。
視細胞とは

視細胞は網膜に存在する神経細胞である。網膜に達した光は、視細胞の外節とよばれる部分で受容される。
視細胞には桿体細胞と錐体細胞の2種類がある。
桿体細胞と錐体細胞
桿体細胞
桿体細胞(rod cell)は、明るさや暗さを感知する視細胞である。桿体細胞は色素としてロドプシンをもち、この色素が光を受容する。
桿体細胞は色の識別はできない一方、外節が大きいため、わずかな光を感じることができる。したがって、暗所で活発になる。
錐体細胞
錐体細胞(cone cell)は網膜の中止部にある視細胞である。感度が低いため暗所では働かず、明所で機能する。
光の三原色に関わる三種類の細胞(赤錐体、緑錐体、青錐体)をもち、色や形を感知する。色覚異常は、少なくとも一種類の錐体細胞が欠損している障害である。
錐体細胞には、ロドプシンとよく似た物質であるフォトプシンが存在する。フォトプシンもオプシンとレチナールから構成されている。
ロドプシン
ロドプシンとは
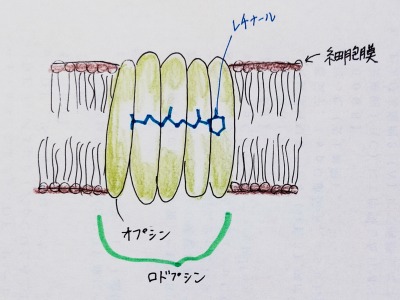
ロドプシン(rhodopsin)は視覚に関わる膜タンパク質であり、光を認識し、電気信号として視神経に伝える役割を担う。このように、光を認識して働くタンパク質を「光受容タンパク質」という。
ロドプシンは、オプシンというタンパク質とレチナールからなる複合体である。

画像引用元:Wikipedia
オプシンはαヘリックスが膜を7回貫通した構造をしており、その中に光を吸収する色素分子としてレチナールが結合している。
光によるロドプシンの変化

オプシンと結合しているレチナールは、シス型(11-cis-レチナール)の構造をしているが、光を受けるとトランス型(all-trans-レチナール)になる。
シス型
レチナールがシス型のとき、オプシンとレチナールが結合している「不活性状態」である。
トランス型
レチナールがシス型からトランス型に変化すると、構造の変化によりオプシンとレチナールの結合が維持できず、解離する。
この構造の変化によって桿体細胞は「活性状態」となる。その刺激は視神経を通って脳に伝わり「視覚」が生じる。
光受容のメカニズム

①光シグナルが電気シグナルに変わる
外界からの光シグナルを視物質(オプシンやレチナール)が受容する。
レチナールは、光を吸収すると構造が変化してシス型からトランス型となり、オプシンから解離する。この立体構造の変化によって生じたメタロドプシンⅡが、Gタンパク質を活性化させる。
Gタンパク質はトランスデューシン(Gt)ともよばれる。
トランスデューシンのサブユニットがGDPからGTPに置換されることで、cGMP(環状グアノシン―リン酸)が分解され、GMP(グアノシン―リン酸)となる。
その結果、cGMPによって活性化されて開口していた「cGMP依存性Na+チャネル」が閉じ、細胞内へのNa+やCa2+の流入が停止する。
暗所では、細胞外のNa+とCa2+がcGMP依存性Na+チャネルを通じて細胞内に流入する。その結果、視細胞の膜電位は「脱分極」状態となっている。
一方、光を受容してcGMP依存性Na+チャネルが閉じると負電荷が増加するため、視細胞の膜電位は速やかに「過分極」状態となる。
この過程で、電位が生じることにより、光シグナルが電気シグナルに変換される。
過分極と脱分極
神経細胞の内外には電位差(電圧差)がある。通常時は、細胞内が細胞外に比べて-70mV~-80mV程度である。これを過分極(抑制)という。興奮していないときの膜電位を「静止膜電位」という。
この電位差が-40mV~-20mVにまで上昇する状態を脱分極(興奮)という。脱分極すると、膜電位依存性ナトリウムチャネルが活性化して神経細胞が活性化される。興奮するときの膜電位の変化を「活動電位」という。
視細胞の特異性
視細胞の場合は通常の神経細胞と異なり、暗所では脱分極(興奮)し、光を吸収するとcGMP依存性Na+チャネルが閉じて過分極(抑制)する。
②電気シグナルの伝達
光によって視細胞が過分極すると、シナプス終末部からの神経伝達物質の放出が減少または停止する。その結果、シナプス後細胞の双極細胞の膜電位を変化させて電気シグナルが伝達される。
双極細胞からの信号は網膜の他の細胞に伝達され、最終的に視神経を通じて視覚情報が脳へ送られる。その情報が脳の視覚野で解釈され、「見える」ようになる。
眼疾患
失明をきたす眼疾患である網膜色素変性症と加齢黄斑変性症についてまとめる。
網膜色素変性症
網膜色素変性症は遺伝性疾患であり、その原因遺伝子として100種以上が報告されている。
通常、桿体細胞から障害されるために、暗所でものがみえにくい「夜盲」の症状が最初に現れる。進行すると視野が狭くなり、錐体細胞も障害されて最終的に失明に至ることがある。
加齢黄斑変性症
加齢黄斑変性症は、老化によって網膜の黄斑部に出血やむくみが生じ、視力が低下する眼疾患である。
黄斑部は網膜の中心部であり、カメラではフィルムの役割をもつ。黄斑部に老廃物が沈着して網膜に異変をきたす。
オプトジェネティクス
オプトジェネティクス(optogenetics)は光遺伝学ともよばれ、光に応答するタンパク質を細胞に導入することによって細胞の機能を操作することをいう。
オプトジェネティクスにより目的細胞の活動を操作することで、行動にどのような影響を与えるのかを調べることができる。
例えば、以下の行動のメカニズムを研究するために用いられる
- 記憶形成
- 攻撃性
- 睡眠
- 不安
- 抑うつ
- 薬物依存
マウスのニューロンの活動を操作
オプトジェネティクスの例として、チャネルロドプシン遺伝子を用いたマウスの実験がある。
チャネルロドプシンの遺伝子を、マウスの視床下部のニューロンに導入する。視床下部は、自室神経系の中枢となる重要な部位であり、体温調節やホルモン分泌、怒りや不安などの情動行動の調節などを行う。
そして、脳に細い光ファイバーを埋め込んだマウスの視床下部を青色光で照射すると、マウスは攻撃性を示すようになる。
これは、光の照射によってチャネルロドプシンが開き、チャネルロドプシンをもつニューロンが脱分極して刺激されたと考えられる。
光の照射を止めると、マウスの攻撃性はなくなる。
チャネルロドプシン

チャネルロドプシンは光合成緑藻類にみられる光依存性イオンチャネルである。このイオンチャネルは、青色光に反応して陽イオン(Na+やCa2+)を細胞内に流入させる。
陽イオンが細胞内に流入することによって膜電位差が生じ、光エネルギーが電気信号として神経細胞に伝わる。

画像引用元:Wikipedia
例えば、チャネルロドプシンをもつクラミドモナスの場合、膜電位差が鞭毛に伝わることで鞭毛の動かし方を変え、クラミドモナスは光の方向へと移動し、光合成を行う。
微生物ロドプシン
ロドプシンは動物ロドプシンと微生物ロドプシンに大きく分類することができる。これらのうち、視機能を回復させるための研究において注目されているのが、微生物ロドプシンである。
微生物ロドプシンが視機能を回復させる
動物ロドプシンの場合、主な機能は光を受容するとGタンパク質にシグナルを伝達することである。
一方、微生物ロドプシンは光を受容するとイオンを運ぶなど様々な機能をもつ。
タンパク質分子として単独で光を感受し、膜電位を変化させることができるロドプシンは、光遺伝学の主要ツールとしての期待が高い。
例えば、藻類のクラミドモナス由来のチャネルロドプシン2(ChR2)をコードする遺伝子を、残存する網膜神経節細胞に導入することによって光受容能を与え、視機能を回復させる研究がおこなわれている。
参照
- オプトジェネティクス(Optogenetics) ~光遺伝学~
http://kandori.web.nitech.ac.jp/research/optogenetics/ - 網膜の不思議
https://www.santen.co.jp/ja/healthcare/eye/eyecare/wonders/retina.jsp - 脊椎動物の視覚と視細胞
http://gabriel.ess.sci.osaka-u.ac.jp/html/hisatomi/phototransduction/PR-jp.html -
錐体細胞
https://human-cell.com/archives/suitai.html - なぜ見えるのか?目の仕組みを解説
https://www.hirabari-ganka.com/news-column/187/ - 光を信号へと変換するタンパク質の新型ヘリオロドプシンを発見
https://www.jst.go.jp/pr/announce/20180621/index.html - CHLAMYDOMONAS FLAGELLA AND PHOTOBEHAVIORS
http://www.res.titech.ac.jp/~junkan/Hisabori_HomePage/chlamydomonas.html - チャネルロドプシン
http://web.cc.iwate-u.ac.jp/~htomita/vis-neurosci/optogenetics.html - 藻類の光感受性タンパク質が、視力を失われた患者の視機能を回復する希望!
https://modia.chitose-bio.com/articles/57/ - ロドプシンの作動メカニズム
https://seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2019.910472/data/index.html





